
LIFE IN THE SEA
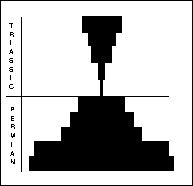
Fusulinid foraminifera disappeared completely, although other foram groups suffered much lower levels of extinction. Palaeozoic corals (Rugosa and Tabulata) also vanished. Stenolaemate bryozoans and articulate brachiopods suffered near-complete extinction. The extant echinoderm groups all experienced severe bottlenecks at this time: only two lineages of crinoids and echinoids made it into the Mesozoic. Several echinoderm groups (e.g. Blastoidea) suffered complete extinction.
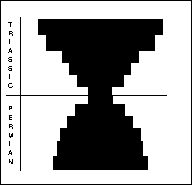
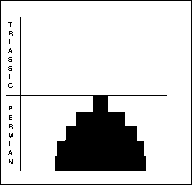 Fusilinid foraminifera (right) were unicellular animals, ranging in size from 0.1-8mm. Benthonic or planktonic forms with a considerable disparity in morphology. Peaked in the Early Carboniferous, already in prolonged decline throughout Permian, and the final 10% died out during the PTME. The non-fusilinid foraminifera (far right) included five suborders. The different suborders were affected differently by the extinction: the Allogromiina are poorly known, the Textulariina lost a third of genera; Miololina lost half of the genera; Lagenina and Involutina showed dramatic increase in post extinction radiation.
Fusilinid foraminifera (right) were unicellular animals, ranging in size from 0.1-8mm. Benthonic or planktonic forms with a considerable disparity in morphology. Peaked in the Early Carboniferous, already in prolonged decline throughout Permian, and the final 10% died out during the PTME. The non-fusilinid foraminifera (far right) included five suborders. The different suborders were affected differently by the extinction: the Allogromiina are poorly known, the Textulariina lost a third of genera; Miololina lost half of the genera; Lagenina and Involutina showed dramatic increase in post extinction radiation.

 Sponges (Porifera), the basal animal group (left), composed of an internal skeleton of spicules of calcite or silica, and forming reefs. There were 13 Late Permian genera, of which four survived into the Triassic.
Sponges (Porifera), the basal animal group (left), composed of an internal skeleton of spicules of calcite or silica, and forming reefs. There were 13 Late Permian genera, of which four survived into the Triassic.
Tabulate corals (below, left) were colonial in form, and had been dwindling through the Permian, and the final seven species were lost at the PTME. The rugose corals (below, next) were solitary and radially symmetrical, comprising solitary and colonial types. Many species were present throughout the Permian, and all became extinct at the PTME.
Bryozoans (below, right) are colonial organisms, with a skeleton of chitin or calcium carbonate, forming small colonies, in fan-shaped, stick-like or lobate forms. Bryozoans suffered greatly in a prolonged decline of species through the Permian: 128 genera fell to 28 in the Late Permian, and five families with vastly reduced diversity survived through to the Triassic, but became extinct soon after.
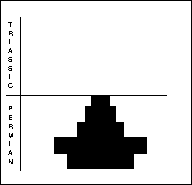
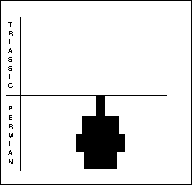


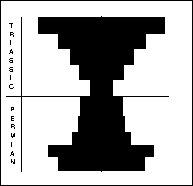
 Brachiopods (right) are bivalved, benthic animals with unequal valves, each of which is symmetrical, composed of chitin or calcium carbonate. The larger, pedicle valve has an aperture through which a muscular stalk protrudes to anchor the animal to the substrate. The valves are opened and closed by adductor muscles, with filter feeding internal organs supported by internal structures. Brachiopods were numerically the most important fossils in Late Permian beds. Sixty genera decreased to 10 through the PTME.
Brachiopods (right) are bivalved, benthic animals with unequal valves, each of which is symmetrical, composed of chitin or calcium carbonate. The larger, pedicle valve has an aperture through which a muscular stalk protrudes to anchor the animal to the substrate. The valves are opened and closed by adductor muscles, with filter feeding internal organs supported by internal structures. Brachiopods were numerically the most important fossils in Late Permian beds. Sixty genera decreased to 10 through the PTME.
Bivalves (below, left) have two equivalent valves, generally made from aragonite. The valves are opened and closed by muscles, which allows filter feeding. They declined from 50 genera (112 species) at the start of the Late Permian to only 9 genera at the P-T boundary. Gastropods (below, next) comprise a single coiled shell. Some 32 genera were seen at the Middle-Late Permian boundary, and dwindled and seemingly disappeared at the P-T boundary. Tiny microgastropods radiated in the earliest Triassic.
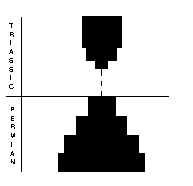
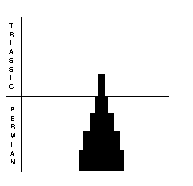
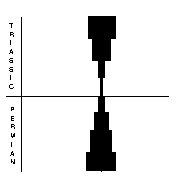
Orthocerid cephalopods (above, right) had a straight, to slightly coiled shell usually made of aragonite. They possessed a soft-bodied “foot” that was modified to a ring of tentacles around the mouth. The group was already in dramatic decline in the Early Permian, and continued to decline throughout. Only two genera and one genus survived into the Triassic and these quickly became extinct. The nautiloids (above, far right) had straight, curved or coiled shells, composed mostly of aragonite. The were well represented in the Late Permian, with 17 species in 11 genera. Eight genera survived into the Triassic and radiated from there.

 The ammonoids (left) were marine molluscs with straight, curved or coiled shells. They were filled with gas or light fluid and could adjust their buoyancy. They include three orders: the Goniatitida were the bulk of Permian ammonoids, but were in decline through the Permian, and all but one died out at the PTME; the Prolecanitida were a small order which was declining throughout Permian, and only just survived across the P-T boundary, before dying out completely in the early Triassic; and the Ceratitida diversified in the Permian, before being decimated by the extinction event, and then reappearing in the Early Triassic, and radiating to become the dominant Triassic forms.
The ammonoids (left) were marine molluscs with straight, curved or coiled shells. They were filled with gas or light fluid and could adjust their buoyancy. They include three orders: the Goniatitida were the bulk of Permian ammonoids, but were in decline through the Permian, and all but one died out at the PTME; the Prolecanitida were a small order which was declining throughout Permian, and only just survived across the P-T boundary, before dying out completely in the early Triassic; and the Ceratitida diversified in the Permian, before being decimated by the extinction event, and then reappearing in the Early Triassic, and radiating to become the dominant Triassic forms.
Echinoids (below, far left), or sea urchins, are entirely marine organisms with a test made from calcareous plates, each of which is a single crystal of calcite. Emerging from pores on the perforate rows are tube feet, which are connected to an internal water vascular system responsible for locomotion and respiration. The group was not diverse either before or after the PTME, so the event had little effect on them. Blastoids (below, left) were also marine, with a test made from calcareous plates, each of which is a single crystal of calcite. Blastoids consist of a stalk surmounted by a calyx which contains 13 plates arranged in three rows. There were small numbers of taxa in the Permian and they became extinct before the Permo-Triassic boundary. The crinoids (below, right) were entirely marine organisms with tests made from calcareous plates. Crinoids consist of a stem surmounted by a calyx of 2 or 3 rows of plates, bearing 5 pinnate arms. These arms collect small food particles which are passed into the mouth. The stem is made from individual ossicles, which make up the majority of crinoid remains found. Crinoids suffered substantial extinction in the PTME, and one genus, Isocrinus (below, far right ) survived and was the root of the recovery of Triassic crinoids.
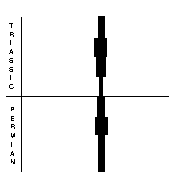

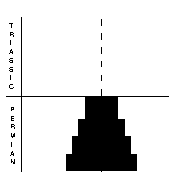

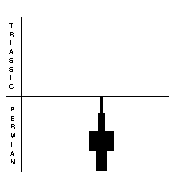
 Trilobites (right) were typical marine arthropods of the Palaeozoic. Their exoskeletons were composed of chitin or calcium carbonate, and they had jointed limbs. They had already dwindled to only two genera in the Late Permian, and disappeared by the time of the PTME. Ostracods (far right) were also arthropods, comprising paired valves, and generally small, measuring 0.5-10 mm. There were some 35 Permian genera, with 87 species. Most of these disappeared during the PTME, and relatively few survived into the Triassic, when diversity rose again to 88 genera.
Trilobites (right) were typical marine arthropods of the Palaeozoic. Their exoskeletons were composed of chitin or calcium carbonate, and they had jointed limbs. They had already dwindled to only two genera in the Late Permian, and disappeared by the time of the PTME. Ostracods (far right) were also arthropods, comprising paired valves, and generally small, measuring 0.5-10 mm. There were some 35 Permian genera, with 87 species. Most of these disappeared during the PTME, and relatively few survived into the Triassic, when diversity rose again to 88 genera.

 The conodonts (left) were microscopic, 0.05-2 mm phosphatic structures. Conodonts occur as different forms, all tooth-like in shape, and they were parts of feeding apparatuses of basal vertebrates. They were in decline through the Permian and Triassic.
The conodonts (left) were microscopic, 0.05-2 mm phosphatic structures. Conodonts occur as different forms, all tooth-like in shape, and they were parts of feeding apparatuses of basal vertebrates. They were in decline through the Permian and Triassic.
LIFE ON LAND
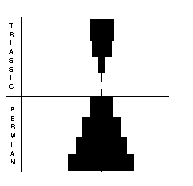
The Permo-Triassic boundary seemingly did not see a dramatic extinction of plants, rather a gradual and diachronous transition from palaeophytic to mesophytic floras. The changeover took place in different areas and at different times over a period of over 25 million years. The transition first occurred in North America and Western Europe, where it was completed by the beginning of the Late Permian, while in Asia and Australasia, palaeophytic forms continued into the Early Triassic. Knoll (1984) stated that “wherever fossiliferous sequences are complete enough, demonstrably paleophytic and mesophytic assemblages are separated by intermediate floras that contain elements of both groups”.
 These evolutionary changes were associated with increasing temperatures and increasing aridity through the Permian. Plants and their seeds and spores had to adapt to retain water to prevent dehydration and death. Water-loving plants such as lycopods and sphenopsids became very small, as shrubs, with ferns and seed ferns being the dominant species.The palaeophytic/ mesophytic transition has been recognised for over 100 years (Weiss, 1877) as a great evolutionary step in the Late Permian, marked by the arrival of gymnosperms, including conifers and gingkos.
These evolutionary changes were associated with increasing temperatures and increasing aridity through the Permian. Plants and their seeds and spores had to adapt to retain water to prevent dehydration and death. Water-loving plants such as lycopods and sphenopsids became very small, as shrubs, with ferns and seed ferns being the dominant species.The palaeophytic/ mesophytic transition has been recognised for over 100 years (Weiss, 1877) as a great evolutionary step in the Late Permian, marked by the arrival of gymnosperms, including conifers and gingkos.
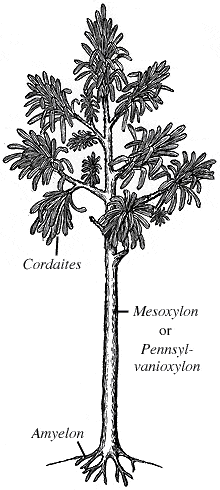
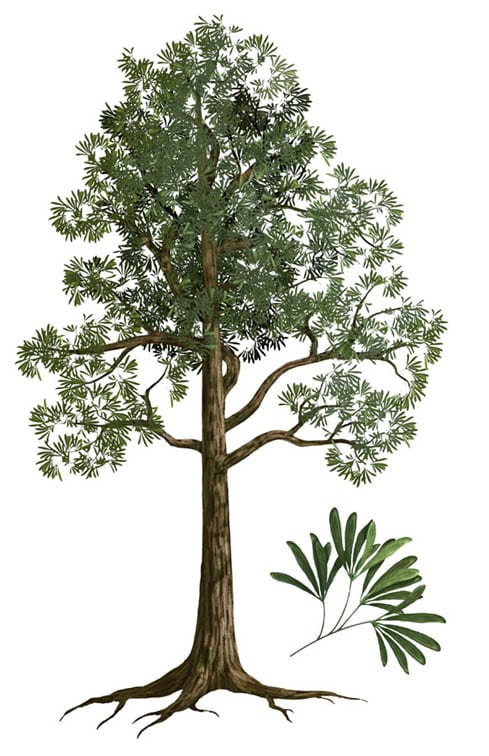
Species numbers of plants did decline through the PTME, with the extinction of the cordaitaleans at the end of the Permian. These were mangrove-like swamp trees (left). Further, the Glossopteris floras (right), which were dominant in Gondwana in the Permian, were replaced by the seed fern Dicroidium as the climate dried.
The evolution of terrestrial tetrapods through the PTME is described here.