
Comparing Russia and South Africa
Our most striking finding has been that the high diversity and complex latest Permian terrestrial ecosystems in Russia were volatile in terms of generic and familial turnover, but that when these ecosystems were largely destroyed by the PT crisis the volatility disappeared, and recovery from low diversity was a slow process (see Figure below), with longer survivorship of genera and families and less turnover. Within the 15 Ma post-event window, full recovery of the ecosystems had not taken place.
 Turnover of tetrapod families through the Late Permian and Early Triassic in the South Urals Basin, Russia. Rates of origination and extinction are percentage metrics based on all taxa (including Lazarus taxa, but excluding singleton families – families known from a single species or single locality) known from a time bin. Stratigraphic units are the successive svitas of the Upper Permian (1, Osinovskaya; 2, Belebey; 3, Bolshekinelskaya; 4, Amanakskaya; 5, Malokinelskaya/ Vyakovskaya; 6, Kutulukskaya/ Kulchomovskaya), Lower Triassic (7, Kopanskaya; 8, Staritskaya; 9, Kzylsaiskaya; 10, Gostevskaya; 11, Petropavlovskaya) and Middle Triassic (12, Donguz; 13, Bukobay). Binomial 95% confidence intervals are shown for the percentage extinction metrics (confidence intervals are of similar magnitude for the percentage origination metrics, but are omitted for clarity). (From Benton et al. 2004.)
Turnover of tetrapod families through the Late Permian and Early Triassic in the South Urals Basin, Russia. Rates of origination and extinction are percentage metrics based on all taxa (including Lazarus taxa, but excluding singleton families – families known from a single species or single locality) known from a time bin. Stratigraphic units are the successive svitas of the Upper Permian (1, Osinovskaya; 2, Belebey; 3, Bolshekinelskaya; 4, Amanakskaya; 5, Malokinelskaya/ Vyakovskaya; 6, Kutulukskaya/ Kulchomovskaya), Lower Triassic (7, Kopanskaya; 8, Staritskaya; 9, Kzylsaiskaya; 10, Gostevskaya; 11, Petropavlovskaya) and Middle Triassic (12, Donguz; 13, Bukobay). Binomial 95% confidence intervals are shown for the percentage extinction metrics (confidence intervals are of similar magnitude for the percentage origination metrics, but are omitted for clarity). (From Benton et al. 2004.)
This contrasts with conclusions from elsewhere. Smith & Botha (2005) reported a 69% generic extinction rate, based on collection of 225 specimens, across the PTB in South Africa. Only four genera, Lystrosaurus, Tetracynodon, Moschorhinus and Ictidosuchoides, survived the PTB. But, within 37 m of the boundary, Smith & Botha (2005) reported a total of ten genera, compared to 13 just below the PTB, and interpreted this to mean that recovery was relatively fast, really within a few hundred thousand years of the mass extinction event.
What is recovery?
Of course, it is important to define what is meant by ‘recovery’. To some, recovery would mean simply the recovery of species numbers in faunas – when post-extinction generic diversity of faunas matches the pre-extinction diversity. In the Russian faunas, this was achieved in the Early Triassic, at the time of the Gostevskaya Sand Petropavlovskaya Svitas, when there were again ten or eleven families, as in the terminal Permian Vyatskian Svita. However, that probably does not represent full recovery in the sense of recovery of the species numbers and ecological roles in faunas. In other words, during Gostevskaya and Petropalovskaya Svita times, the families present did not include any small insectivores or large herbivores. In that sense, the ecosystems were incomplete and large herbivores at least did not come on the scene until latest Middle Triassic or early Late Triassic, some 20 myr after the PT mass extinction.
A third definition of recovery could be the recovery of higher-taxon diversity on a global scale. On this count, terrestrial tetrapod faunas world-wide had not recovered the Late Permian global patterns until the Late Triassic and the time of origin of the dinosaurs and other groups, again some 20 myr after the PT mass extinction. For marine families, the global recovery period was even longer, extending into the Early Cretaceous, when global marine family numbers recovered their Late Permian levels, some 125 myr after the PT mass extinction. In this regard, the global recovery of marine family numbers was set back further by the end-Triassic mass extinction.
So, the Russian evidence suggests a rather slow ecosystem recovery lasting for more than 20 myr, whereas Smith & Botha (2005) proposed a rapid recovery in South Africa over a time span of less than 1 myr. The difference in findings reflects partly a difference in definition, whether recovery indicates overall ecosystem recovery (Russian example) or species diversity in faunas (South Africa).
Sampling might play a role, but so too might the sedimentary environments represented. The apparent relative abundance of amphibians in the Early Triassic of both South Africa and Russia has long been noted (e.g. Milner 1990) and could reflect, to some extent, a bias relating to the kinds of rock facies preserved. Ironically, while the earliest Triassic was a time of increasing aridity in Russia (Newell et al. 1999) and South Africa (Smith & Botha, 2005), the wet-adapted amphibians predominate in some localities. Smith & Botha (2005) noted specific evidence in the Karoo of drought-induced deaths of Lystrosaurus and other taxa and, in both regions, overall drought might have been associated with rare monsoonal rainfall that produced massive floods and coarse river deposits, as well as skeletons of short-lived amphibians that flourished during the weeks of wet conditions.
A possible bias in comparing the two regions is the relative abundance of the famous dicynodont Lystrosaurus in South Africa, and its complete absence from the Orenburg region (although the genus is known elsewhere in the Russian Early Triassic (Surkov et al. 2005). Further, although truly large herbivores are not known in the Triassic units of the Karoo, as in Russia, there were insect-eating reptiles among the cynodonts that are not known in the Early and Mid Triassic of Orenburg, and only rarely from elsewhere in the Russian successions (Battail & Surkov 2000). The absence of dicynodonts and cynodonts in the Early Triassic of the Russian South Urals area, and their relative rarity elsewhere in Russia, might indicate a palaeobiogeographical difference between South Africa and Russia, but further consideration of the distribution of sedimentary facies and sampling controls must be considered.
Among other terrestrial groups, plants seem to show a slow recovery, lasting until the end of the Middle Triassic (Grauvogel-Stamm & Ash 2005). Indeed, in several parts of the world, including Russia and South Africa, the recovery did not really start until the end of the Early Triassic: after the extinction of Permian floras, the lycopsid Pleuromeiaproliferated worldwide. Then, conifers re-established themselves in the early Anisian, and new groups – cycadophytes and pteridosperms – appeared in the late Anisian. The timing of floral recovery world-wide is more in line with the slow recovery of tetrapod faunas seen in Russia than with the rather more rapid recovery of tetrapods suggested from South Africa.
Ecological aspects of recovery
In an alternative approach to understanding recovery, Sahney & Benton (2008) looked at changes in ‘alpha diversity’, that is number of species in an ecocystem/ geological formation, through the Permian and Triassic (see Figure below). Disaster taxa, such as Lystrosaurus, insinuated themselves into almost every corner of the sparsely populated landscape in the earliest Triassic, and a quick taxonomic recovery apparently occurred on a global scale.
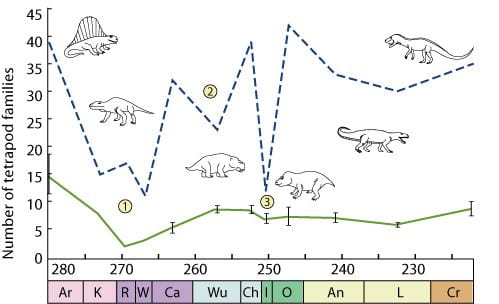 Figure 1. Global diversity (dashed line) and mean alpha diversity (solid line) of Permo-Triassic tetrapod families. Extinctions are labelled as 1, Olson’s extinction; 2, end- Guadalupian extinction; and 3, end-Permian extinction. Geological stages (Gradstein et al. 2004) are as follows: Ar, Artinskian; K, Kungurian; R, Roadian; W, Wordian; Ca, Capitanian; Wu, Wuchiapingian; Ch, Changhsingian; I, Induan;O, Olenekian; An, Anisian; L, Ladinian; Cr, Carnian.
Figure 1. Global diversity (dashed line) and mean alpha diversity (solid line) of Permo-Triassic tetrapod families. Extinctions are labelled as 1, Olson’s extinction; 2, end- Guadalupian extinction; and 3, end-Permian extinction. Geological stages (Gradstein et al. 2004) are as follows: Ar, Artinskian; K, Kungurian; R, Roadian; W, Wordian; Ca, Capitanian; Wu, Wuchiapingian; Ch, Changhsingian; I, Induan;O, Olenekian; An, Anisian; L, Ladinian; Cr, Carnian.
However, close study of ecosystem evolution shows that true ecological recovery was slower. After the end-Guadalupian event, faunas began rebuilding complex trophic structures and refilling guilds, but were hit again by the end-Permian event. Taxonomic diversity at the alpha (community) level did not recover to pre-extinction levels; it reached only a low plateau after each pulse and continued low into the Late Triassic. Our data showed that though there was an initial rise in cosmopolitanism after the extinction pulses, large drops subsequently occurred and, counter-intuitively, a surprisingly low level of cosmopolitanism was sustained through the Early and Middle Triassic. It was not until the great diversity of the Late Triassic, which included dinosaurs, pterosaurs, crocodilians, rauisuchids, aetosaurs, rhynchosaurs, trilophosaurs, sphenodonts, amphibians and mammals, some 30 Myr after the end-Permian event, that terrestrial tetrapod community diversity was restored.
Posture shift among tetrapods across the PTB
In a further contribution, Kubo & Benton (2009) noted an astonishing observation: Permian tetrapods were sprawlers, and Triassic tetrapods were upright in posture and gait. It seems that the mass extinction had an astonishing effect on both the diapsid and synapsid lines of tetrapods, whose posture evolved in parallel – a kind of ancient ‘arms race’ between predators and prey. Read more here.
References
- Benton, M.J., Tverdokhlebov, V.P. and Surkov, M.V. 2004. Ecosystem remodelling among vertebrates at the Permian-Triassic boundary in Russia. Nature 432, 97-100 (doi:10.1038/nature02950). pdf Download the original of Figure 1 in colour as a pdf. Download the Excel data file here.
- Gradstein, F.M., Ogg, J.G. & Smith, A.G. 2004. A geologic time scale 2004. Cambridge University Press, Cambridge.
- Grauvogel-Stamm, L. & Ash, S.R. 2005. Recovery of the Triassic land flora from the end-Permian life crisis. Comptes Rendus Palévol, 4, 593-608.
- Kubo, T. and Benton, M.J. 2009. Tetrapod postural shift estimated from Permian and Triassic trackways. Palaeontology 52, 1029-1037. pdf. Interpretive web site.
- Milner, A.R. 1990. The radiations of temnospondyl amphibians. In (Taylor, P.D. & Larwood, G.P.; eds) Major evolutionary radiations. Systematics Association Special Volume. Clarendon Press, Oxford, 321-349.
- Newell, A.J., Tverdokhlebov, V.P., and Benton, M.J. 1999. Interplay of tectonics and climate on a transverse fluvial system, Upper Permian, southern Uralian foreland basin, Russia. Sedimentary Geology 127, 11-29. pdf
- Sahney, S. and Benton, M.J. 2008. Recovery from the most profound mass extinction of all time. Proceedings of the Royal Society, Series B 275, 759-765. pdf
- Smith, R.H.M. & Botha, J. 2005. The recovery of terrestrial vertebrate diversity in the South African Karoo Basin after the end-Permian extinction. Comptes Rendus Palévol, 4, 623-636.
- Surkov, M.V., Kalandadze, N.N. & Benton, M.J. 2005. Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia. Journal of Vertebrate Paleontology, 25, 402-413.